Media optimization in the RoboLector platform for enhanced protein production using C. glutamicum
Introduction
In the biotechnological industry, medium and process optimization is highly demanded. Optimization processes are often associated with time-consuming steps in order to achieve the highest possible throughput. However, the amount of work should always be minimized while maximizing the information content at the same time. The BioLector, a microbioreactor system, combines the scalable BioLector technology of monitoring biomass, pH, DO, and diverse fluorescences together with a microfluidic chip for feeding and pH-control. In combination with the RoboLector platform it is a perfect tool for performing media preparations and automated high-throughput cultivation processes for applying design of experiment (DoE) conditions. The system is based on a standard microtiter plate (MTP) format and operates with non-invasive optical measurement during the fermentation process. This technology can be used to implement process control via pneumatically controlled microchannels for cultivation well-individual pH control and fed-batch cultivations.
In the research project1 described in this application note, we are presenting a media optimization of the minimal medium for Corynebacterium glutamicum (CGXII medium) using the RoboLector platform and BioLector microbioreactor. The media optimization in the DoE is focused on increasing the cutinase activity in C. glutamicum. The organism is cultivated in batch and fed-batch processes using the BioLector microbioreactor in order to investigate the influence of the media components and feeding rates on the growthrelated secretory protein production of cutinase. In this context, we further demonstrate a successfully performed microbial cultivation within the high cell density range in micro liter scale.
Methods
Media components: The original standard minimal medium for C. glutamicum (CGXII-standard) consists of 42 g/L 3-(Nmorpholino)-propane sulfonic acid (MOPS), 22 g/L glucose, 20 g/L ammonium sulfate, 5 g/L urea, 1 g/L phosphate buffer, 13.25 mg/L calcium chloride, 0.25 mg/L magnesium sulfate heptahydrate, 10 mg/L iron sulfate heptahydrate, 10 mg/L manganese sulfate monohydrate, 1 mg/L zinc sulfate heptahydrate, 0.2 mg/L copper sulfate, 0.02 mg/L nickel chloride hexahydrate, 0.2 mg/L biotin, 30 mg/L protocatechuic acid (PCA), 50 mg/L kanamycin and 59.6 mg/L isopropyl β-d-1-thiogalactopyranoside (IPTG).
The BioLector-adapted CGXII medium (CGXII-BLP) contains 5.5 g/L 3-(N-morpholino)- propane sulfonic acid (MOPS), no ammonium sulfate and 6 mg PCA. The rest of the components remain in the same concentration as listed for the CGXII-standard medium. With the CGXII-BLP medium the media optimization is initiated.
Sensitivity analysis: A sensitivity analysis is an essential tool to figure out the optimization potential of the certain media and to identify the media components having a negative or positive effect on the microorganisms with regard to both the growth and the protein production. This experimental series is considered as preparation step for the actual optimization in the DoE. Here, the effect of every media component is regarded individually. To that end, the individual component concentrations of the medium to be optimized were halved and then doubled in the next experiment. Only one component in the medium was changed at a time in order to subsequently observe the growth behaviour in the batch process using the BioLector. At the end of the test series the media components with the highest positive influence on the cutinase activity are determined: magnesium sulfate, iron sulfate and manganese sulfate.
Media optimization with DoE-Builder: Three media components in different concentration combinations are investigated which have had the most positive effect on secretory cutinase production in the previous sensitivity analysis: The concentrations of magnesium sulfate, iron sulfate and manganese sulfate are varied by the DoE Builder. The concentration range of magnesium sulfate is between 0 and 500 mg/L, for iron sulfate between 0 and 20 mg/L and for manganese sulfate between 0 and 20 mg/L. The other components and concentrations remained constant. As a control the medium without concentration changes (CGXII-BLP) is used. Finally, the adaptation of the models to the cutinase activity offers 15 experiments with suitable concentration combinations. A total of two optimization steps are performed.
Media preparation using RoboLector: For the automated media preparation the pipetting robot is used. This method leads to an increase in throughput during media production. Method validation is not necessary as there is no significant difference in microbial growth between the manual and automated approach2. The automated method is used exclusively for the media preparation of the batch tests, since the composition of the optimized medium remains constant in the fed-batch tests.
Cultivation strain: In this project C. glutamicum (ATCC 13032) pEKEx2-NprE-Cutinase is cultivated in CG XII medium. This strain was kindly provided by Institute of Bio- and Geoscience (IBG-1), Forschungszentrum Jülich GmbH, Germany. The cutinase originates from Fusarium solanini pisi. The signal peptide derived from Bacillus subtilis provides the secretion via Sec-Pathway.[9]
Cultivation conditions: For the first preculture, 100 µL of a C. glutamicum pEKEx2-NprE-Cutinase cryoculture is cultivated overnight in 10 mL Brain Heart Infusion (BHI) + 50 mg/L kanamycin in a 250 mL shake flask at 30 °C and 250 rpm with a shaking diameter of 5 cm. A second preculture is performed in 10 mL of CGXII-standard medium in a 250 mL shake flask at 30 °C and 250 rpm with a shaking diameter of 5 cm overnight.
The batch cultivations of C. glutamicum are performed in a 48 well FlowerPlate (MTP-48- BOH1), incubated in CGXII-optimized medium in the BioLector at 30 °C and 1200 rpm with an initial OD600 of 0.5 and a filling volume of 800 µL per well. The MTP is sealed with a gas permeable and evaporation reducing foil (F-GPR).
The fed-batch cultivations of C. glutamicum are carried out in a microfluidic 32 well FlowerPlate (MTP-MF32-BOH1), incubated in CGXII-optimized medium in the BioLector at 30 °C and 1200 rpm with an initial OD600 of 0.5 and a filling volume of 800 µL per cultivation well, row C-F of the microfluidic FlowerPlate. The MTP is sealed with a gas permeable and evaporation reducing foil (F-GPRSMF32). The pH is controlled to a value of 7. For this, row A of the microfluidic FlowerPlate is filled with 3 M phosphoric acid, 1800 µL in each reservoir well. The glucose feeding is performed with linear feeding profiles with 1 g/(L∙h) + 0.1 µL/h, 2 g/(L∙h) + 0.1 µL/h, 3 g/(L∙h) + 0.1 µL/h and 4 g/(L∙h) + 0.1 µL/h glucose feed. The feeding can be described with equation (1), here for 3 g/(L∙h) + 0.1 µL/h as an example:
f(t) = 5,45 µL/h + 0,1 µL/h (1)
The activation trigger for the linear feeding strategy is set to a setpoint of > 14 h, right after the batch-phase is over. The feeding solution (containing 440 g/L glucose and 100 g/L urea) is filled in row B of the microfluidic FlowerPlate, 1800 µL per reservoir well.
Biomass calibration: For the determination of cell dry weight (CDW) a dilution series of C. glutamicum between OD600 = 0 and OD600 = 37.2 is prepared and measured spectrophotometrically at 600 nm in triplicates.
1 mL of each OD dilution step is centrifuged at 4000 rpm for 5 minutes each in pre-dried (at 80 °C for 24 h) and weighed 1.5 mL reaction tubes. Supernatant is discarded and the cell pellet is resuspended in 1 mL phosphate buffer saline (PBS). The centrifugation step is repeated and the supernatant is discarded again. After drying at 80 °C for 24 h the weight difference is used to calculate the CDW. The backscatter (BS [a.u.]) measurement (for biomass gain 3 and 5) with the same OD600 dilution series was calibrated in a microfluidic FlowerPlate (MTP-MF32-B), with a filling volume of 800 µL at 1200 rpm and 30 °C. The backscattered light values are measured for at least 30 minutes. The backscattered light calibration is leading to the linear regression equation of CDW [g∙L -1 ] and OD [-] described in equation (2) and (3) with R2 > 0.99.
CDW = 0.5009 g∙L -1 ∙a.u.-1 ∙BS – 1.1956 g∙L -1 (2)
OD = 2.0044 a.u.-1 ∙BS – 3.4291 (3)
In figure 1 the biomass (gain 3) calibration curve for the CDW calculation is shown exemplary.

Figure 1 Calibration curve for the CDW in the BioLector
Cell harvest: The cell harvest takes place at the end of the exponential phase or in the early stationary phase when cutinase activity is the highest. For sampling, the BioLector is connected to the RoboLector. With the help of the RoboLector, samples can be collected always at the same time point during the stationary growth phase in each cultivation run. The samples arethen centrifuged and stored cool in the Tecan plate reader until further processing.
Enzyme assay: The lipolytic enzyme activity of cutinase in the culture supernatant is determined by an enzymatic lipase assay3. The test is based on the hydrolysis of p-nitrophenyl-palmitate (pNPP) to p-nitrophenol (pNP). The basic pH of the reaction buffer leads to deprotonation of pNP to 4-nitrophenolate. Absorption is detected in the plate reader (Safire 2, Tecan) at a wavelength of 410 nm.
Results
Influence of the PCA concentration: For cultivation processes in the BioLector the PCA concentration needs to be reduced to 6 mg/L in order to avoid interfering effects in the performance of the pH optodes. In previous studies, a better adapted medium (CGXII-BLP) was developed for better results in the BioLector4 . Moreover, the ammonium component has been removed. The PCA component cannot be removed completely from the medium since PCA is essential for the growth phase and reproducibility of the biological replicates4. However, the negative influence of the PCA component on the pH optodes in the FlowerPlate may cause that the pH value measured by the BioLector differs from the pH value measured offline. Consequently, the modified CGXII-BLP medium with less PCA must be used for further experiments, to ensure that the deviation between online and offline pH values are as small as possible.
Sensitivity analysis: For the assessment of a possible improvement in the growth of C. glutamicum and an increased cutinase activity the scattered light signal of the biomass was converted to the optical density using the biomass calibration on the BioLector. Ten different media components of the CGXII-BLP medium areinvestigated and evaluated in 38 different sets with six biological replicates each. In the first part of this sensitivity analysis, described in figure 2 and figure 3, ten different media components with respect to growth and cutinase activity are compared. In the second part of this sensitivity analysis (see figure 4 and figure 5) seven different media components without the influence of zinc sulfate and copper sulfate are investigated. Figure 2 shows the OD and cutinase activity after reduction of the concentrations by half. The values for the optical density in all cases are below the control values. In addition, the results show that C. glutamicum reacts differently to the concentration of the corresponding component in each sample being reduced by half. For example, a reduction in trace elements such as iron sulfate leads to lower protein secretion.

Figure 2 Sensitivity analysis with copper sulfate and zinc sulfate, concentrations of the media components are reduced by half.
In comparison, a concentration doubling (shown in figure 3) of iron sulfate leading to a higher cutinase activity. Here it is 14.5 % higher than with the concentration reduction. Trace elements containing metal ions influence the cell wall structure and thus have a positive effect on secretion performance5. Furthermore, the doubling of copper sulfate and zinc sulfate concentration can have a negative influence on the growth of C. glutamicum and on the cutinase activity as well.

Figure 3 Sensitivity analysis with copper sulfate and zinc sulfate, concentrations of the media components are doubled.
The second sensitivity analysis is performed with a modified medium free of copper sulfate and zinc sulfate for their presence have a negative effect on microbial growth and the cutinase activity in C. glutamicum. First, the concentrations of the remaining media components are reduced (figure 4) and doubled as well (figure 5). The concentration increase of urea is leading to a higher growth, however, the cutinase activity is 13 % higher when the urea concentration is reduced. In general, the removal of the components copper sulfate and zinc sulfate promotes higher cutinase activities. It is shown that both halving and doubling the concentrations of manganese sulfate, iron sulfate and magnesium sulfate have a positive effect on C. glutamicum when both copper sulfate and zinc sulfate are removed compared to the control where copper sulfate and zinc sulfate are contained.

Figure 4 Sensitivity analysis without copper sulfate and zinc sulfate, concentrations of the media components are reduced by half.
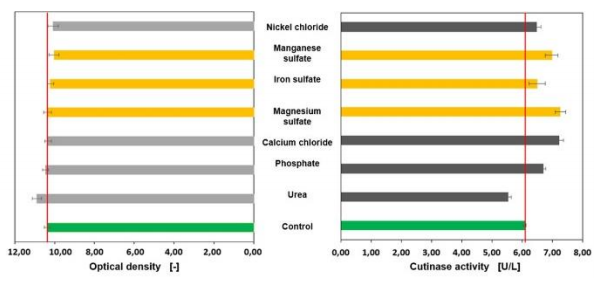
Figure 5 Sensitivity analysis without copper sulfate and zinc sulfate, concentrations of the media components are doubled.
The media components magnesium sulfate, iron sulfate and manganese sulfate have the highest impact on the performance of C. glutamicum. The selection of these three media components was not only based on the results of an increased target size in the CGXII-BLP recipe medium, but is also due to the fact that metal ions not only have an influence on the net charge of the target protein, which alone has an influence on the secretion performance, but also on the proteins of the secretion system8. The percentage changes in relation to the results of the tests both with and without copper sulfate and zinc sulfate were determined by doubling the concentration of the respective media components as shown in table 1. When doubling magnesium sulfate, an increase of 0.19 % can be achieved in the medium without copper and zinc sulfate and 4.48 % for manganese sulfate. The optical density with a doubling of the iron sulfate component is 5.71 % lower than in the medium supplemented with copper and zinc sulfate.
Table 1 Relative change of target values in media w/o zinc and copper sulfate with doubled media component concentration.
|
OD[-] |
Cutinase activity[U/L] |
|
| Magnesium sulfate | + 0,19 % | + 8,18 % |
| Iron sulfate | - 5,71 % | + 5,69 % |
| Manganese sulfate | + 4,48 % | + 22,67 % |
Media optimization using DOE builder: Based on the results of the sensitivity analysis for the media optimization the media components magnesium sulfate, iron sulfate and manganese sulfate are further investigated in two optimization steps using DoE-builder Modde® (Umetrics) to plan the experiments and verify the concentrations. Media optimization is performed with the modified CGXII-BLP medium without zinc sulfate and copper sulfate since both components have a negative impact on C. glutamicum regarding the cutinase production. For the first optimization step 15 experiments are designed. The combinations for the first optimization step are listed in table 2. For magnesium sulfate the concentration range is chosen between 0 and 500 mg/L, for iron sulfate between 0 and 20 mg/L and for manganese sulfate 0 to 20 mg/L. The remaining concentration of the components of the CGXII-BLP medium stay constant. In each optimization step a control experiment is done with the modified CGXII-BLP medium without zinc sulfate and copper sulfate. Hence, 15 different experiments with four biological replicates each are performed in the FlowerPlate using the BioLector.
Table 2 Combination of the component concentrations of the CGXII-BLP medium for the first optimization step
| Experiment | Magnesium sulfate [g/L] | Iron sulfate [g/L] | Manganese sulfate [g/L] |
| N1 | 0 | 0 | 0 |
| N2 | 0.5 | 0 | 0 |
| N3 | 0 | 20 | 0 |
| N4 | 0.5 | 20 | 0 |
| N5 | 0 | 0 | 20 |
| N6 | 0.5 | 0 | 20 |
| N7 | 0 | 20 | 20 |
| N8 | 0.5 | 20 | 20 |
| N9 | 0 | 10 | 10 |
| N10 | 0.5 | 10 | 10 |
| N11 | 0.25 | 0 | 10 |
| N12 | 0.25 | 20 | 10 |
| N13 | 0.25 | 10 | 0 |
| N14 | 0.25 | 10 | 20 |
| Control | 0.25 | 10 | 10 |
In the first optimization step, 15 experiments with 4 biological replicates each are compared. In figure 6 the determined values for the OD and cutinase activity for each experiment are shown.

Figure 6 Determined values for the optical density and the cutinase activity in the first optimization step
The results regarding the optical density in the first optimization step show in particular that growth depends on the magnesium sulfate component (see figure 6: N1, N3, N5, N7 and N9). For example, in case of missing magnesium sulfate component the growth decreases by 80 % compared to the control experiment. Thus, magnesium sulfate is the determining criterion regarding an increase in the cutinase activity. The experiments N4 and N13 show that the absence of a manganese sulfate component has no relevant influence on the growth and cutinase production of C. glutamicum. This phenomenon is shown especially by the approaches N8 and N10, which lead to the highest cutinase concentrations. For N10 the iron sulfate and manganese sulfate components are available in the original concentration and for N8 the concentrations for latter components are doubled. The cutinase activity is 8.02 U/L for N8 and 7.96 U/L for N10. In comparison, between control and N8, optical density increased by 9.33 % and cutinase activity increased by 39.23 %. Thus, magnesium sulfate is the decisive factor for an increased cutinase production. In the next step, the determined cutinase activities are entered into the software to give input for the second optimization step. Using the multiple linear regression (MLR) method in the DoE. The 4D plotsdemonstrate the dependence of the cutinase activity on the three variable parameters shown in figure 7 for 0 mg/L manganese sulfate, in figure 8 for 10 mg/L manganese sulfate and in figure 9 for 20 mg/L manganese sulfate. The plots show the ranges of the highest expected activity colored in red. In the second optimization step, the optimizer tool provides the following factor settings and theoretical targets for improving cutinase activity.

Figure 7 4D plot of the varying concentrations of iron sulfate and magnesium sulfate with 0 mg/L manganese sulfate
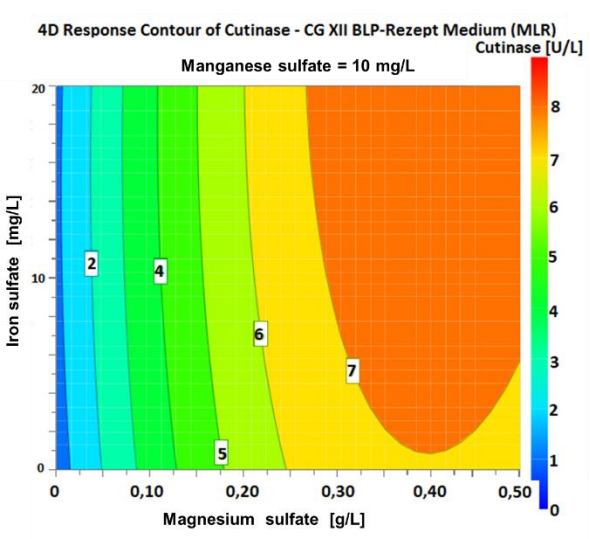
Figure 8 4D plot of the varying concentrations of iron sulfate and magnesium sulfate with 10 mg/L manganese sulfate
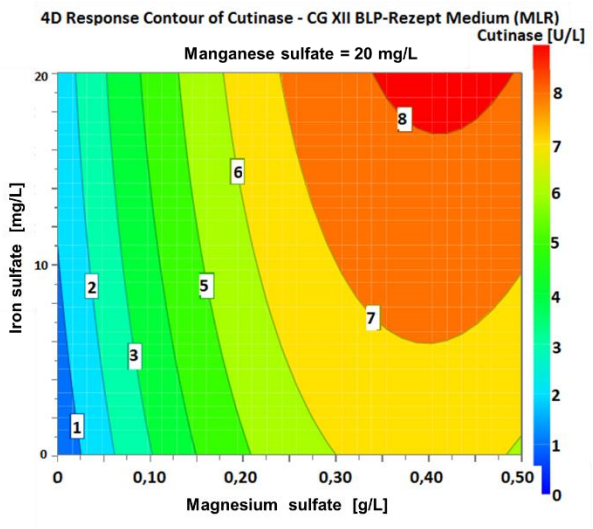
Figure 9 4D plot of the varying concentrations of iron sulfate and magnesium sulfate with 20 mg/L manganese sulfate
Based on the results of the first optimization step, 24 theoretically improved experiments are calculated and performed in a batch process in the FlowerPlate using the BioLector. The combinations for the second optimization step are listed in table 3. The results regarding the optical density and cutinase activity are shown in figure 10. Here, the control approaches N1 and N 13 are mean averaged and indicated as “control”
Table 3 Combination of the component concentrations of the CGXII-BLP medium for the second optimization step
| Experiment | Magnesium sulfate [g/L] | Iron sulfate [g/L] | Manganese sulfate [g/L] |
| N1* | 0,25 | 10 | 10 |
| N2 | 0,375 | 0 | 0 |
| N3 | 0,415 | 20 | 20 |
| N4 | 0,421 | 20 | 14,83 |
| N5 | 0,40 | 20 | 20 |
| N6 | 0,425 | 20 | 20 |
| N7 | 0,41 | 20 | 5 |
| N8 | 0,38 | 19,6 | 20 |
| N9 | 0,398 | 18,2 | 20 |
| N10 | 0,45 | 19 | 20 |
| N11 | 0,50 | 25 | 25 |
| N12 | 0,40 | 25 | 20 |
| N13* | 0,25 | 10 | 10 |
| N14 | 0,50 | 25 | 20 |
| N15 | 0,30 | 20 | 20 |
| N16 | 0,55 | 20 | 20 |
| N17 | 0,60 | 20 | 20 |
| N18 | 0,38 | 15 | 20 |
| N19 | 0,35 | 17 | 20 |
| N20 | 0,50 | 19 | 20 |
| N21 | 0,40 | 22 | 20 |
| N22 | 0,55 | 22 | 20 |
| N23 | 0,50 | 25 | 25 |
| N24 | 0,25 | 25 | 20 |
| * N1 and N13 are control approaches | |||
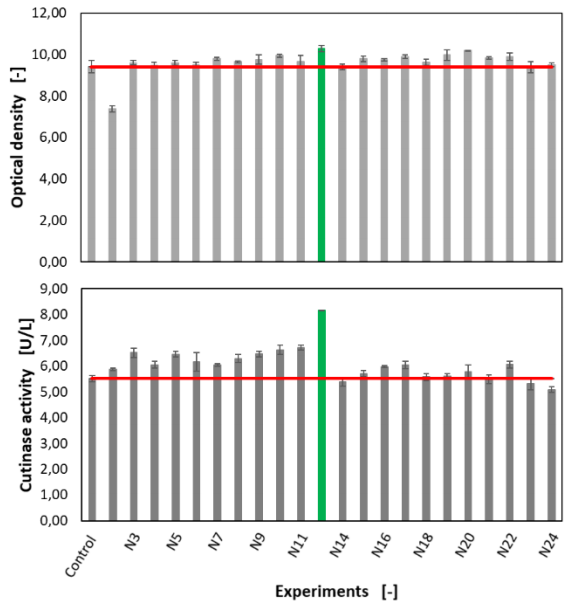
Figure 10 Determined values for the optical density and the cutinase activity in the second optimization step
The difference in the optical density is negligible in all experiments except for experiment N2 since in all approaches apart from that one all media components are present and the organism grows comparably well. Moreover, in the second optimization step, the concentrations between the different approaches differ only minimally. In contrast, the secretory protein production and thus the cutinase activity shows significant differences between the individual batches as shown in figure 10. Here, the media composition of approach N12 shows a clear improvement in terms of cutinase activity compared to the control experiment and the best result from the first optimization step (see N8 in figure 6). In comparison, to the control approach, the optical density in N12 is increased by 9.46 % and the cutinase activity by 47.82 %. Compared to N8 from the first optimization step, the optical density is improved by 4.57 % and the cutinase activity by1.75 %. The final optimized medium (CGXIIoptimized) developed in this test series contains 400 mg/L magnesium sulfate, 25 mg/L iron sulfate and 20 mg/L manganese sulfate. This corresponds to a concentration increase of 60 % for magnesium sulphate, 150 % for iron sulphate and100 % for manganese sulphate compared to the CGXII-BLP medium. The increase in concentration of the metal ions shows the importance of these in combination with magnesium sulfate for secretory protein secretion. One reason for this is that metal ions have an influence on protein secretion through the bacterial cell wall. Bacterial cell walls are able to bind cations due to a high negative charge density. This property enables the folding of secretory proteins 6,7.
Comparison of different feeding rates: The successful development of the optimized CGXII medium using the DoE discloses the possibility of conducting experiments with different feeding rates in the BioLector to find out the ideal fed-batch conditions for a high protein production. The feeding rate of the substrate solution is an important parameter since it has a direct influence on the microorganism in terms of biomass growth and cutinase formation. For this purpose, the feeding phase of a two-phase process is investigated. In the first phase, C. glutamicum grows in a batch culture with a final substrate concentration of 5 g/L glucose in the optimized CGXII medium. Subsequently, a linear glucose feed is started. The culture is treated with an oxygen concentration of 35 % in the head space over the entire cultivation time. Moreover, the pH is controlled to pH 7.0 using 3 M phosphoric acid.In total, four different feed rates of the linear feeding strategy are adjusted to check and compare the influence of the substrate supply on the secretory cutinase production. An exemplary cultivation process in the optimized CGXII medium is shown in figure 11.

Figure 11 Fed-batch cultivation of C. glutamicum in the optimized CGXII medium with a linear feed rate of 5.45 µL/h + 0,1 µL/h in the BioLector® Pro
Here the growth curve and the dissolved oxygen are presented with a linear feeding profile with a feed rate of 3 g/L∙h + 0.1 µL/h. The feed is starting after 14 h and thus the fed-batch phase is initiated. At this time point no oxygen limitation is observable. After completion of the batch phase, when the optical density is 9.88, the linear feed is started. Subsequently, the biomass signal increases continuously to an optical density of 17.13 for about 21 h. After 21 h, the cells start to grow exponentially. At the same time the dissolved oxygen starts to decrease. After 28.5 h an oxygen limitation is visible and is manifested by a slowed down biomass growth after 31 h. After 37.5 h, the optical density rises to OD = 110 and flattens after 45 h. This behavior at the late stageof cultivation indicates a limitation of C. glutamicum either due to the lack of essential trace elements or oxygen limitation. In addition, the DO signal increases strongly after 49.5 h showing the growth of the culture is stopped at this time point. In figure 12 the cutinase activities of C. glutamicum for different linear feeding rates in the BioLector are shown.

Figure 12 Comparison of the cutinase activities of C. glutamicum at different linear feed rates with an increase of 0.1 µL/h
Here, the four different feeding rates 1 g/L∙h + 0.1 µL/h, 2 g/L∙h + 0.1 µL/h, 3 g/L∙h + 0.1 µL/h and 4 g/L∙h + 0.1 µL/h are compared. Each approach with eight biological replicates of the individual feed rates has been cultivated under the same environmental conditions in BioLector in the optimized CGXII medium. The results of the linear feed procedure with C. glutamicum show that the highest cutinase activity is achieved at a feed rate of 2 g/L∙h + 0.1 µL/h with of 45.64 U/L. This result is 11.70 % higher than the cutinase activity at a feed rate of 1 g/L∙h, 10.80 % higher than the cutinase activity at 3 g/L∙h and 21.93 % higher than the cutinase activity at 4 g/L∙h. The cutinase activities and the corresponding productivities for each feed rate are presented in table 4. Here it is shown that the highest productivity is achieved at a feed rate of 2 g/L∙h + 0.1 µL/h with 0.88 U/L∙h.
Table 4 Overview of the cutinase activities and productivities at different linear feed rates in the BioLector
| Feed rate | Cutinase activity [U/L] | Productivity [U/L∙h] |
| 1 g/L∙h + 0.1 µL/h | 40.30 | 0.77 |
| 2 g/L∙h + 0.1 µL/h | 45.64 | 0.88 |
| 3 g/L∙h + 0.1 µL/h | 40.71 | 0.78 |
| 4 g/L∙h + 0.1 µL/h | 35.63 | 0.68 |
In figure 13 the optical density for the same feeding rates are shown.

Figure 13 Comparison of the optical densities of C. glutamicum at different linear feed rates with an increase of 0.1 µL/h
At a feed rate of 1 g/L∙h + 0.1 µL/h, the lowest optical density of 56.88 is reached. For 2 g/L∙h + 0.1 µL/h the optical density is 106.95, for 3 g/L∙h + 0.1 µL it is 115.40 and for 4 g/L∙h + 0.1 µL/h an optical density of 110.54 can be achieved. In summary it can be seen that a feed rate of 1 g/L∙h + 0.1 µL/h is not sufficient enough for the entire cultivation period, since the optical density only reaches a value of 56.88, which is 46.82 % lower than the OD achieved with a feed rate of 2 g/L∙h. With regards to the cutinase activity with 40.30 U/L it is 11.70 % below the activity at a feed rate of 2 g/L∙h. Furthermore, the results of the feed rates 3 g/L∙h and 4 g/L∙h clearly show: the higher the linear feed rate, the higher the decrease in cutinase production of C. glutamicum due to higher oxygen limitation.
Conclusion
This application note shows an exemplary media optimization with the RoboLector platform in order to achieve an enhanced protein production in C. glutamicum. Furthermore, the optimized medium is used in fed-batch cultivations in the BioLector for the purpose to find out the ideal feeding conditions for a high cell density and cutinase activity in C. glutamicum. Here it is shown that the RoboLector is an ideal device for carrying out media optimization in DoE environment and validation experiments in fedbatch mode.
References
[1] Meyer, F. (2019): Medien- und Prozessoptimierung von C. glutamicum mit Hilfe des RoboLector, Master’s Thesis, RWTH-Aachen University.
[2] Rohe, P.; et al. (2012): An automated workflow for enhancing microbial bioprocess optimization on a novel microbioreactor platform. In: Microbial cell factories 11: 144.
[3] Winkler, U.; Stuckmann, M. (1979): Glycogen, hyaluronate, and some other polysaccharides greatly enhance the formation of exolipase by Serratia marcescens. In: Journal of Bacteriology 138 (3): 663–670.
[4] Morschett, H.; et al. (2019): Parallelized microscale fed-batch cultivation in onlinemonitored microtiter plates: implications of media composition and feed strategies for process design and performance. In: J Ind Microbiol Biotechnol 46: 1-13.
[5] Kikuchi, Y.; et al. (2008): Production of Chryseobacterium proteolyticum proteinglutaminase using the twin-arginine translocation pathway in Corynebacterium glutamicum. In: Applied microbiology and biotechnology 78 (1): 67–74.
[6] Haddaoui, E.; et al. (1999): Bacillus subtilis alpha-amylase. The rate limiting step of secretion is growth phase-independent. In: FEMS microbiology letters 173 (1): 127-131.
[7] Leloup, L.; et al. (1997): Characterization of the rate-limiting step of the secretion of Bacillus subtilis alpha-amylase overproduced during the exponential phase of growth. In: Microbiology (Reading, England) 143 (10): 3295–3303.
[8] Thwaite, J.; et al. (2002): Optimization of the cell wall microenvironment allows increased production of recombinant Bacillus anthracis protective antigen from B. subtilis. In: Applied and environmental microbiology 68 (1): 227–234.
[9] Hemmerich, J.; et al. (2018): Combinatorial impact of Sec signal peptides from Bacillus subtilis and bioprocess conditions on heterologous cutinase secretion by Corynebacterium glutamicum. In: Biotechnology and Bioengineering 116 (3): 644-655.
The data shown in this Application Note was generated on BioLector II and BioLector Pro microbioreactors. The BioLector model shown in this Application Note is the latest model of the series, a BioLector XT microbioreactor.
