Exosome-Depleted FBS Using Beckman Coulter Centrifugation: The cost-effective, Consistent choice
Content Type: Application Note
Chad Schwartz Ph.D. | Beckman Coulter, Inc., Indianapolis, IN 46268
Abstract & Introduction
Exosomes are small microvesicles, derived from the late endosome, most often described in the literature to be less than 120 nm, released by all cell types, and proven to be involved in cancer metastasis1-3. Exosome characterization and analysis comprise a fast, evolving research area even though their biological function has yet to be completely elucidated. Exosomes contain proteins, lipids, and microRNA capable of regulating an assortment of target genes. Recent studies have suggested that exosomes can serve as biomarkers for future clinical and diagnostic use in not only cancer but many other human diseases4. Exciting new findings have implicated exosomes in cardiovascular diseases5-7, autoimmune syndromes8, and neurodegenerative disorders such as Alzheimer’s9 and Parkinson’s10 disease, in addition to infectious diseases such as tuberculosis11, diphtheria12, and even HIV13.
Much of the research involving exosomes uses a cell culture platform, although exosomes are routinely isolated from other bodily fluids such as plasma, serum, urine, and breast milk. In cell culture, fetal bovine serum (FBS) is usually incorporated into media, despite FBS containing extremely high levels of innate bovine extracellular vesicles which complicate downstream analyses. There has recently been a call for the importance of depleting cell culture media of contaminating bovine exosomes14 within the research community. In order to deplete FBS of bovine exosomes, researchers routinely turn to ultracentrifugation, due to the simplicity and efficiency of the process. In this process, large volumes of FBS can easily be centrifuged at high speeds to eliminate native extracellular vesicles typically in an overnight spin. However, there are also commercially available products which have pre-conditioned the media to be depleted of exosomes and other microvesicles. The process here is proprietary and is dependent on the manufacturer but is often very costly compared to source FBS. Here, we will explain the process for “home-brewing” exosome-depleted FBS by ultracentrifugation and compare the cell viability and media depletion percentage from two separate cell lines after treatment with different media types. Suggested ways to isolate exosomes of interest from cell culture with an example centrifugation protocol will also be discussed.
Materials & Methods
Culture media was prepared in three different ways.
- Source media: 50 mL of standard HI-FBS (Gibco) was added to 450 mL of both MEM (Gibco) for Jurkat cells and RPMI 1640 (Gibco) for HCT 116 cells. The media was then supplemented with 10 mM HEPES and 100 U/mL Penicillin-Streptavidin (Gibco).
- Ultracentrifuged media: 500 mL of standard HI-FBS was added equally to 6 Beckman Coulter Ultra-Clear 94 mL centrifuge tubes (part #: 345777) with an adapter and then placed in a Beckman Coulter Type 45 Ti rotor and spun at 120,000 x g, 18 hours, 4˚C in a Beckman Coulter Optima XPN ultracentrifuge. The supernatant of each tube was recovered and aliquoted to 50 mL and stored in the -20˚C freezer for future use. 50 mL of the centrifugally-depleted FBS was then added to 450 mL of both MEM and RPMI 1640 media. The media was finally supplemented with 10 mM HEPES and 100 U/mL Penicillin-Streptavidin.
- Commercially-depleted media: 50 mL of Exo-FBS
Exosome-depleted FBS (System Biosciences) was added to both 450 mL of MEM and RPMI 1640. The media was also supplemented with 10 mM HEPES and 100 U/mL Penicillin-Streptavidin.
Frozen stocks of both cell lines were thawed and suspended in the separate buffer types and initially added to 6 well culture plates (Becton Dickinson). Cells were expanded as they reached confluency and added to T-175 flasks (Greiner). Viability was assessed by Beckman Coulter’s Vi-Cell on days 3 and 7 of culture with a passage in between. Briefly, HCT 116 cells were trypsinized, resuspended in appropriate buffer, and centrifuged at 500 x g, 5 min, 20˚C in a Beckman Coulter Allegra X-15 R in an SX4750A rotor. Cells were resuspended again in the appropriate buffer and 1 mL was added to vials and placed in the Vi-Cell for analysis. 1 mL of the suspension Jurkat cells was added directly to the Vi-Cell.
For Nanoparticle Tracking Analysis (NTA) of FBS sources to quantify depletion, 12 mL of each source was filtered through a 0.22 μm filter (Millipore) and added to Beckman Coulter Ultra-Clear centrifuge tube (344059) and placed in an SW 41 Ti rotor and spun at 120,000 x g, 2 hours, 4˚C in a Beckman Coulter Optima XPN. The supernatant was aspirated and the pellet was resuspended in 100 μl 1X PBS pH 7.2 (Gibco). The resulting particles were analyzed on a Nanosight v.2.3 (Malvern Instruments).
Exosomes of interest were isolated from both cell types and all three culture conditions using Beckman Coulter centrifugation and characterized for size by Beckman Coulter’s DelsaMax Core. Figure 1A demonstrates the workflow for isolation. After obtaining highly viable cells, assayed by the Vi-Cell, 40 mL of cell culture was added to 50 mL conical tubes and placed in a SX4750A rotor with 50 mL conical adapters in the Allegra X-15R tabletop centrifuge and spun for 10 min at 750 x g. The supernatant was subsequently recovered, filtered through a 0.45 μm filter, and spun at 2000 x g for 20 min. The supernatant was then centrifuged at 10,000 x g for 30 minutes in the Optima XPN ultracentrifuge equipped with a SW 32 Ti rotor to remove cell debris. Again, the supernatant was recovered, filtered through a 0.22 µm membrane, and spun at 100,000 x g for 90 minutes with an SW 41 Ti rotor in an Optima XPN. This time, the supernatant was aspirated and the pellet was recovered by resuspending in phosphate buffered saline (PBS). The resuspended sample is labeled as crude exosomes and is stable at -20˚C for an extended period of time.
 |
| Figure 1A: Cellular exosome recovery and analysis. A workflow is defined to isolate exosomes of interest by Beckman Coulter centrifugation |
| Gradient Layer | Estimated Density (g/mL) | Volume (mL) | % Iodixanol (0.25M Sucrose; pH 7.5) |
| 1 | 1.223 | 3 | 40 |
| 2 | 1.127 | 3 | 20 |
| 3 | 1.079 | 3 | 10 |
| 4 | 1.054 | 2 | 5 |
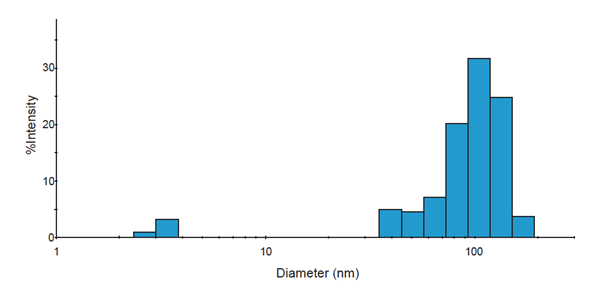 |
| Figure 1C: Representative histogram of DelsaMax dynamic light scattering assay describing exosomes of expected size. |
To further purify the sample, Beckman Coulter’s Biomek 4000 Laboratory Automation Workstation was used to provide a quick, consistent, and reproducible method for layering a centrifugation density gradient with the volumes and density shown in figure 1B. The resuspended crude exosome sample was then layered on top of the gradient and centrifuged at 100,000 x g at 4˚C for 18 hours with an SW 41 Ti rotor and Optima XPN. The Biomek 4000 was employed again to fractionate the gradient after the centrifugation step. 1 mL fractions were collected from the top using liquid level tracking for a total of 13 fractions which were then pelleted using a TLA 120.2 rotor in an Optima Max-XP bench-top centrifuge. The resulting pellet was again resuspended in PBS, analyzed for size, and based on expected density and size of recovered exosomes, fractions 6-9 were combined, pelleted once more using the TLA 120.2 rotor, and finally resuspended in a small volume of PBS. The purified fractions were again analyzed by the Beckman Coulter DelsaMax for size, using a representative trace of Jurkat exosomes (Fig. 1C).
Results
Depletion percentage assayed by NTA
Nanoparticle Tracking Analysis was performed on all FBS sources. The number of traces for centrifugally-depleted media and commercially available media was nominal compared to the source media (Fig. 2) suggesting that both methods were successful in voiding the FBS of contaminating exosomes and other particles.
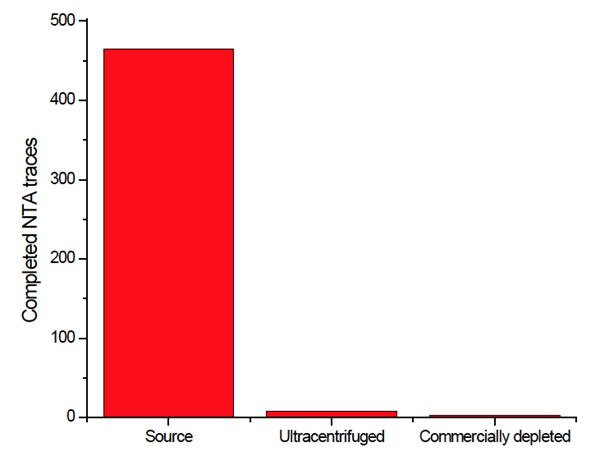 |
| Figure 2: Depletion assay of FBS after treatment. Three FBS sources were centrifuged. The resulting pellet was recovered and analyzed for contaminating exosomes. |
Cell viability of serum type
Cell viability was measured at two different time points prior to exosome isolation for each cell line by Beckman Coulter’s Vi-Cell to understand the effect of depleting exosomes on cell health. Beckman Coulter’s Vi-Cell offers terrific ease-of-use and reproducibility in an automated fashion, relying on high-powered optics and Trypan blue staining for cell count and viability. As noted in Fig. 3, the process of depleting media had little to no effect on the survival of cells.
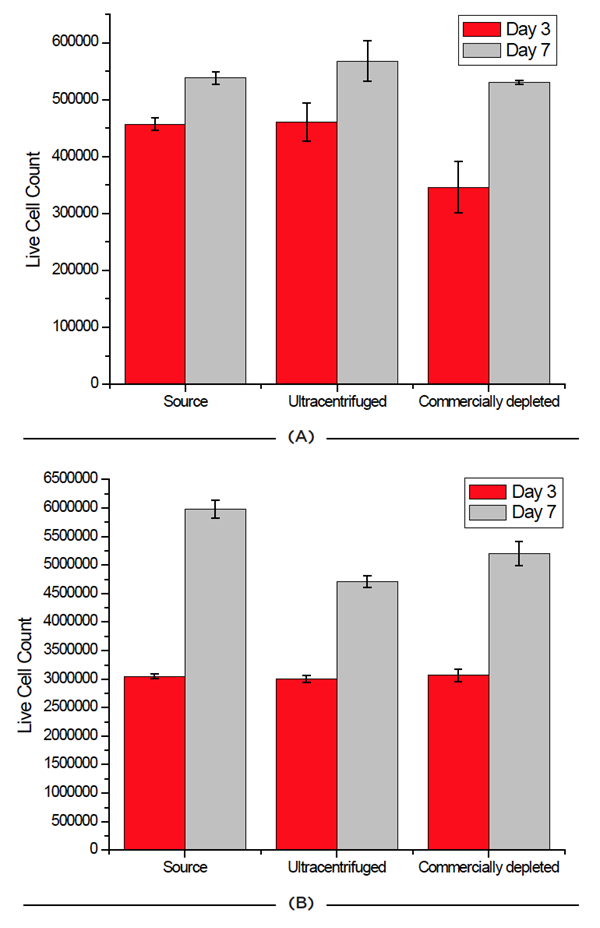 |
| Figure 3: Effect of FBS source on cell viability on two cell lines. Jurkat (A) and HCT 116 (B) cells were grown for seven days and passaged at days 3 and 7. Cell count and viability was measured for all 3 FBS sources on these days. |
Discussion
Exosomes and other extracellular vesicles are present in high concentrations in fetal bovine serum, FBS, a standard supplement to nearly all cell culture conditions. As the field of exosome research continues to expand, it is becoming increasingly important to standardize methods for FBS depletion in experiments utilizing cell culture. Commercially available depleted FBS is currently more than double the cost than other heat-inactivated, highly-pure FBS at most distributors. To ease the pain associated with high experimental costs, an alternative solution has been provided with demonstrated results. The centrifugally-depleted FBS retained cell viability during culture, proved low in contamination of exosomes and other particles as probed by NTA, and provided the means to properly purify cellular exosomes by differential centrifugation and density gradient clarification.
References
- 1Vader, P., Breakefield, X.O., Wood, M.J.. Extracellular vesicles: emerging targets for cancer therapy. Trends Mol Med. 2014. 20(7): 385-93.
- El Andaloussi, S., Mager, I., Breakefield, X.O., Wood, M.J.. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013. 12(5): 347-57.
- Simpson, R.J., Lim, J.W., Moritz, R.L., Mathivanan, S.. Exosomes: proteomic insights and diagnostic potential. Expert Rev. Proteomics. 2009. 6(3): 267-83.
- De Toro, J., Herschlik, L., Waldner, C., Mongini, C.. Emerging roles of exosomes in normal and pathological conditions: new insights for diagnosis and therapeutic applications. Front. Immunol. 2015. doi: 10.3389/fimmu.2015.00203.
- Amabile, N., Rautou, P-E, Tedgui, A., Boulanger, C.M.. Microparticles: key protagonists in cardiovascular disorders. Semin Thromb. Hemost. (2010) 36:907–16. doi:10.1055/s-0030-1267044Asdf
- DeJong O.G., Verhaar, M.C., Chen, Y., Vader, P., Gremmels, H., Posthuma, G., et.al. Cellular stress conditions are reflected in the protein and RNA content of endothelial cell-derived exosomes. J Extracell. Vesicles (2012) 1:18396. doi:10.3402/jev.v1i0.18396.
- Waldenström, A., Gennebäck, N., Hellman, U., Ronquist, G.. Cardiomyocyte microvesicles contain DNA/RNA and convey biological messages to target cells. PLoS One (2012) 7:e34653. doi:10.1371/journal.pone.0034653.
- Robbins, P.D., Morelli, A.E.. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol (2014) 14:195–208. doi:10.1038/nri3622.
- Rajendran, L., Honsho,M., Zahn,T.R., Keller,P., Geiger,K.D., Verkade,P., et. al. Alzheimer’s disease beta-amyloid peptides are released in association with exosomes. Proc. Natl. Acad. Sci. USA (2006) 103:11172–7. Doi:10.1073/pnas.0603838103.
- Danzer, K.M., Kranich, L.R., Ruf, W.P., Cagsal-Getkin, O., Winslow, A.R., Zhu,L., et. al. Exosomal cell-to-cell transmission of alphasynuclein oligomers. Mol Neurodegener (2012) 7:42. doi:10.1186/1750-1326-7-42
- Kruh-Garcia, N.A., Wolfe, L.M., Chaisson, L.H.,Worodria, W.O., Nahid P., Schorey J.S., et. al. Detectionof Mycobacteriumtuberculosis peptides in the exosomes of patients with active and latent M. tuberculosis infection using MRM-MS. PLoS One (2014) 9:e103811. doi:10.1371/journal.pone.0103811.
- Colino, J., Snapper, C.M. Exosomes from bone marrow dendritic cells pulsed with diphtheria toxoid preferentially induce type1 antigen-specific IgG responses in naïve recipients in the absence of free antigen. J Immunol (2006) 177:3757–62. doi:10.4049/jimmunol.177.6.3757.
- Gould S.J., Booth, A.M., Hildreth,J.E.K.. The Trojan exosome hypothesis. Proc Natl Acad Sci USA (2003) 100:10592–7. doi:10.1073/pnas.1831413100.
- Shelke, G.V., Lasser, C., Gho, Y.S., Lotvall, J.. Importance of exosome depletion protocols to eliminate functional and RNA-containing extracellular vesicles from fetal bovine serum. J Extracell Vesicles. 2014. Doi:10.3402/jev.v3.24783.
CENT-1124APP09.15-A
